DNA
在特定的环境条件下(pH、离子特性、离子浓度等),DNA链上的功能团可以产生特殊的氢键、离子作用力、疏水作用力以及空间位阻效应等,从而使得DNA分子的各个原子在三维空间里具有了确定的相对位置关系,这称为DNA的空间结构(spatial structure)。
DNA的空间结构可分为二级结构(secondary structure)和高级结构。
一、DNA的二级结构
(一)DNA双螺旋结构
DNA的二级结构是双螺旋结构。有关DNA中四种碱基的Chargaff规则揭示了DNA的碱基之间存在着某种对应的关系,为碱基之间的互补配对关系奠定了基础。
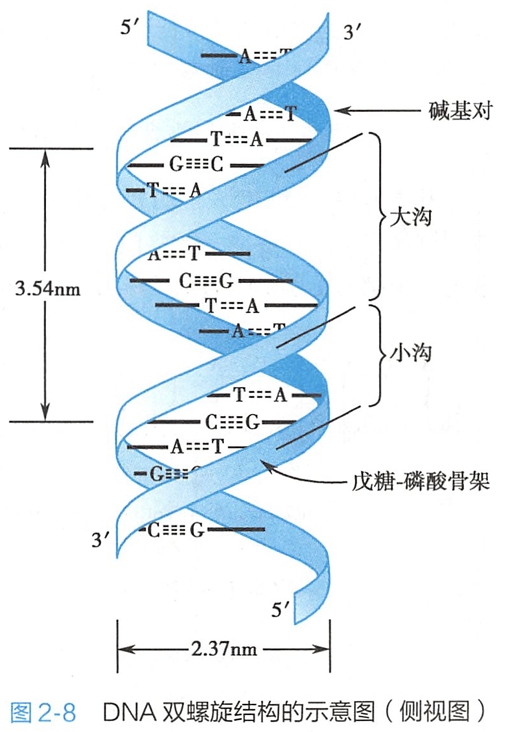
(二)DNA双螺旋结构模型的要点
DNA双螺旋结构具有下列特征:
1.DNA由两条多聚脱氧核苷酸链组成
两条多聚脱氧核苷酸链围绕着同一个螺旋轴形成反平行的右手螺旋(right--handed helix)的结构(图2-8)。两条链中一条链的5'→3'方向是自上而下,而另一条链的5'→3'方向是自下而上,呈现出反向平行(anti-parallel)的特征。DNA双螺旋结构的直径为 2.37nm,螺距为3.54nm。
2.DNA的两条多聚脱氧核苷酸链之间形成了互补碱基对
碱基的化学结构特征决定了两条链之间的特有相互作用方式:一条链上的腺嘌呤与另一条链上的胸腺嘧啶形成了两对氢键;一条链上的鸟嘌呤与另一条链上的胞嘧啶形成了三对氢键(图2-9)。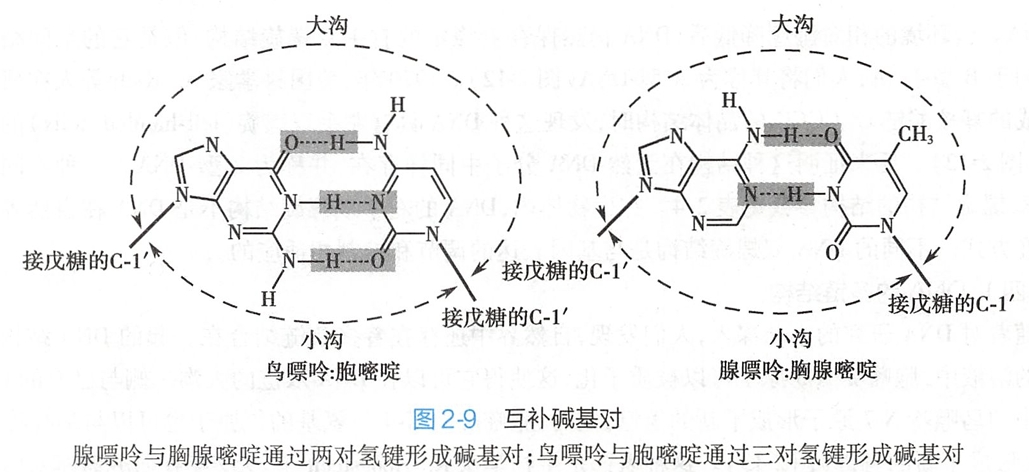 这种特定的碱基之间的作用关系称为互补碱基对(complementary base pair),DNA的两条链则称为互补链(complementary strand)。碱基对平面与双螺旋结构的螺旋轴近乎垂直。平均而言,每一个螺旋有10.5个碱基对,碱基对平面之间的垂直距离为0.34nm。
这种特定的碱基之间的作用关系称为互补碱基对(complementary base pair),DNA的两条链则称为互补链(complementary strand)。碱基对平面与双螺旋结构的螺旋轴近乎垂直。平均而言,每一个螺旋有10.5个碱基对,碱基对平面之间的垂直距离为0.34nm。
3.两条多聚脱氧核苷酸链的亲水性骨架将互补碱基对包埋在DNA双螺旋结构内部
多聚脱氧核苷酸链的脱氧核糖和磷酸基团构成了亲水性骨架(backbone),该骨架位于双螺旋结构的外侧,而疏水性的碱基对包埋在双螺旋结构的内侧(图2-10)。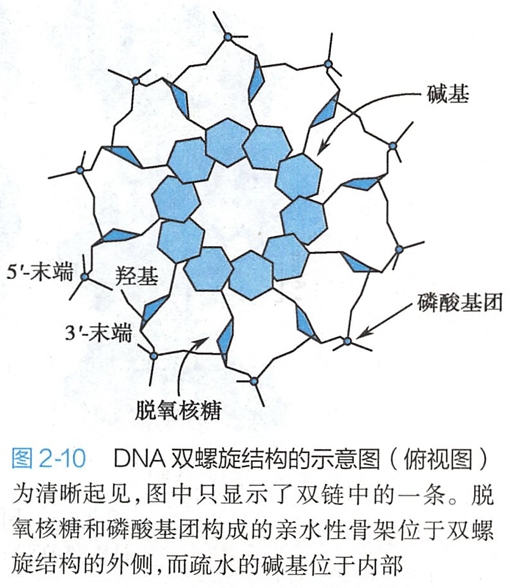
DNA双链的反向平行走向使得碱基对与磷酸骨架的连接呈现非对称性,从而在DNA双螺旋结构的表面上产生一个大沟(major groove)和一个小沟(minor groove)。
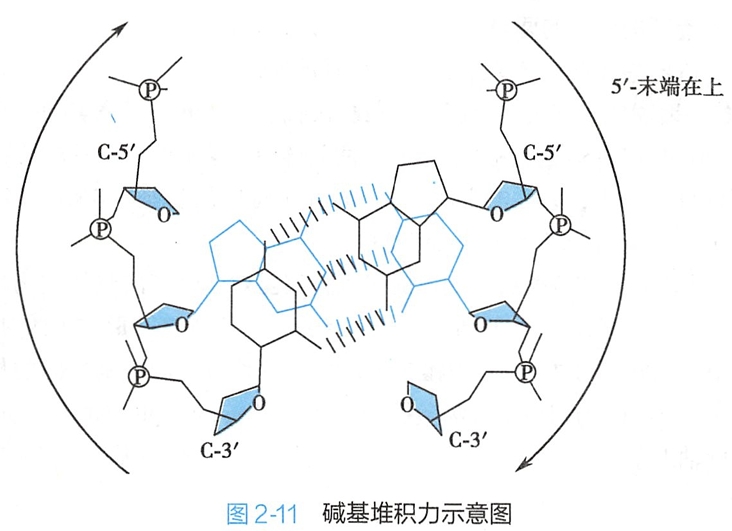
4.两个碱基对平面重叠产生了碱基堆积作用
在 DNA双螺旋结构的旋进过程中,相邻的两个碱基对平面彼此重叠(overlapping),由此产生了疏水性的碱基堆积力(base stacking force)(图2-11)。这种碱基堆积作用十分重要,它和互补链之间碱基对的氢键共同维系着DNA双螺旋结构的稳定。
(三)DNA双螺旋结构的多样性
溶液的离子强度或相对湿度的变化可以使DNA双螺旋结构的沟槽、螺距、旋转角度、碱基对倾角等发生变化。由于历史原因,人们将J.Watson和F.Crick提出的双螺旋结构称为B型-DNA。当环境的相对湿度降低后,DNA仍然保存着稳定的右手双螺旋结构,但是它的空间结构参数不同于B型-DNA,人们将其称为A型-DNA(图2-12)。
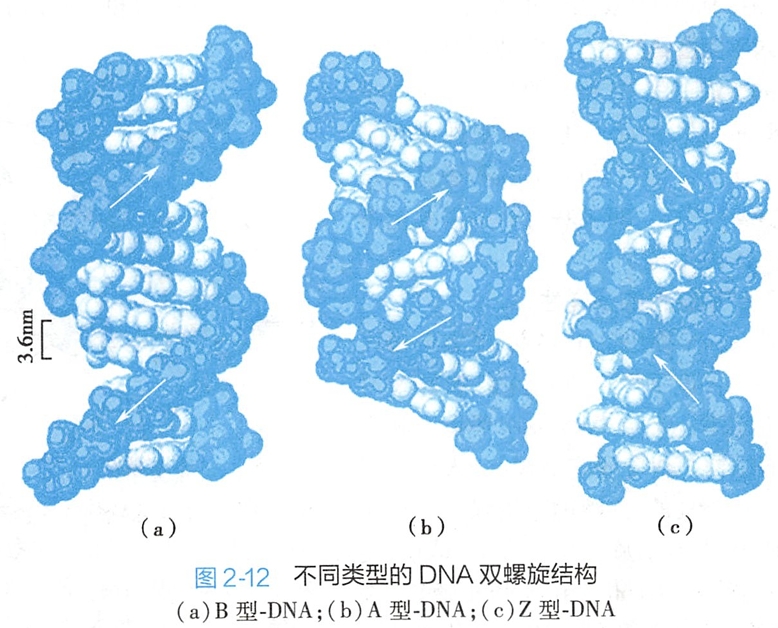
1979年,美国科学家A.Rich等人在研究人工合成的寡核酸链CGCGCG的晶体结构时,发现这种DNA具有左手双螺旋(left-handed helix)的结构特征(图2-12)。后来证明这种结构在天然DNA分子中同样存在,并称为Z型-DNA。
在生物体内,DNA的右手双螺旋结构不是DNA在自然界中唯一存在方式。
(四)DNA的多链结构
自然界中还存在着多条链结合在一起的DNA结构。在酸性的溶液中,胞嘧啶N-3原子可以被质子化,这使得它可以在DNA双链的大沟一侧与已有的GC碱基对中的鸟嘌呤N-7原子形成了新的氢键,同时,胞嘧啶的C-4位氨基的氢原子也可以与鸟嘌呤的C-6位氧形成了新的氢键(图2-13)。
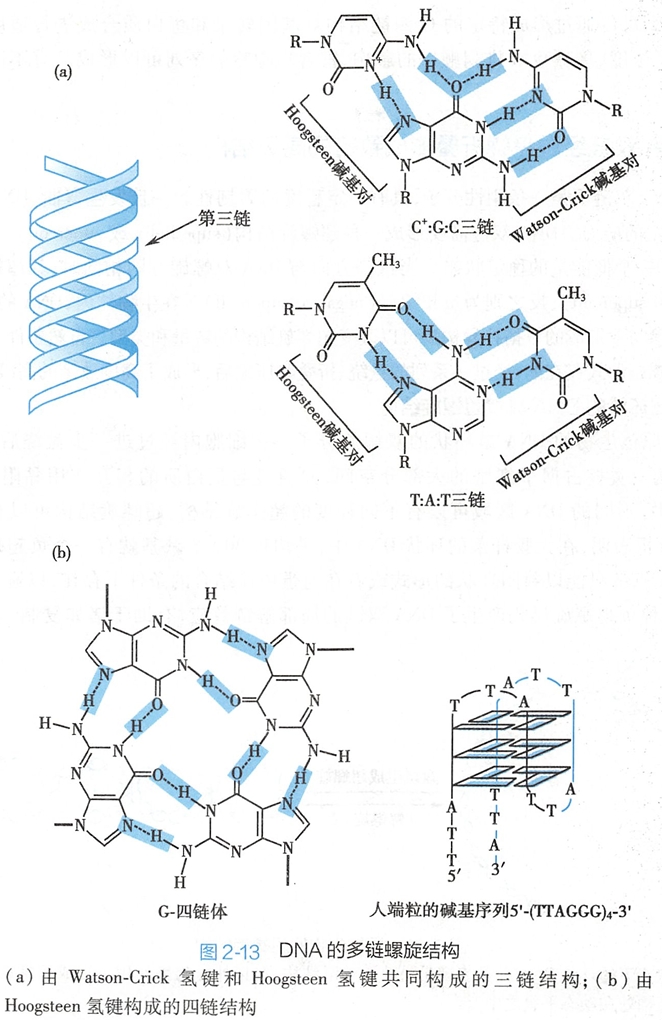
这种氢键是生物学家K.Hoogsteen于1959年在研究碱基对时发现的,故命名为Hoogsteen氢键。Hoogsteen氢键的形成并不破坏原有碱基对中的Watson-Crick氢键,这样就形成了含有三个碱基的C+GC平面,其中GC之间是以Watson-Crick氢键结合,而C+G之间是以 Hoogsteen氢键结合的。
同理,DNA也可以形成TAT的三碱基平面(图2-13)。当DNA双链中一条链的核苷酸序列富含嘌呤时,对应的互补链必然是富含嘧啶,它们形成了正常的DNA双链。如果还有一条富含嘧啶的单链(其序列与富含嘧啶链具有极高的相似度),并且环境条件为酸性时,这条链上的嘧啶就会与双链中的嘌呤形成Hoogsteen氢键,从而生成了DNA的三链结构(triplex)。
人们曾经利用这样的三链结构来尝试着调控基因的表达。根据某些基因的序列特征(例如富含嘌呤的序列),人们设计了富含嘧啶的寡核苷酸链。由于这条寡核苷酸链与这一段双链DNA的序列有着碱基互补关系,它可以嵌入在双链DNA的大沟中形成了三链结构,以此干扰调控因子的结合,影响该基因的复制或转录。
真核生物染色体3'-端是一段高度重复的富含GT的单链,被称为[200 学习/201 细胞生物学/第08章 细胞核/第2节 染色质和染色体/染色质和染色体#(五)端粒|端粒],例如人端粒区的碱基序列是(TTAGGG)。其重复度可达数百乃至上千。作为单链结构的端粒,具有较大的柔韧度,可以自身回折形成一个称为G-四链(G-quadruplex)的特殊结构。这个G-四链结构的核心是由4个鸟嘌呤通过8对Hoogsteen氢键形成的G-平面(tetrad或quartet)(图2-13)。若干个G-平面的堆积使富含鸟嘌呤的重复序列形成了G-四链结构。人们推测这种G-四链结构是用来保护端粒的完整性。
近来,人们还发现某些癌基因的启动子和mRNA的3'-非翻译区都有一些富含鸟嘌呤的序列。这些序列可以通过形成特定的G-四链结构对基因转录和蛋白质合成进行适度的调控。受离子类型、离子浓度、鸟嘌呤G排列顺序的影响,富含鸟嘌呤的序列可以形成具有不同拓扑构象的G-四链体。
二、DNA的高级结构
线性的DNA双链不是一条刚性分子,具有一定程度的柔韧性。一旦发生弯曲,DNA双链就会在其内部产生一定的应力。
DNA双链需要形成一种超螺旋结构(superhelix或supercoil),释放出这些应力使DNA处在一个低能量的稳定状态。
当盘绕方向与DNA双螺旋方同相同时,其超螺旋结构为正超螺旋(positive supercoil);反之则为负超螺旋(negative supercoil)。
在生物体内,DNA的超螺旋结构是在拓扑异构酶参与下形成的。拓扑异构酶可以改变超螺旋结构的数量和类型。自然条件下的DNA双链主要是以负超螺旋形式存在的,经过一系列的盘绕、折叠和压缩后,形成了高度致密的高级结构。
(一)封闭环状的DNA具有超螺旋结构
绝大部分原核生物的DNA是环状的双螺旋分子。在细胞内经过进一步盘绕后,形成了类核(nucleoid)结构。类核占据了细胞的大部分空间,并通过与蛋白质的相互作用黏附在细胞内壁。在细菌DNA中,不同的DNA区域可以有不同程度的超螺旋结构,超螺旋结构可以相互独立存在(图2-14)。
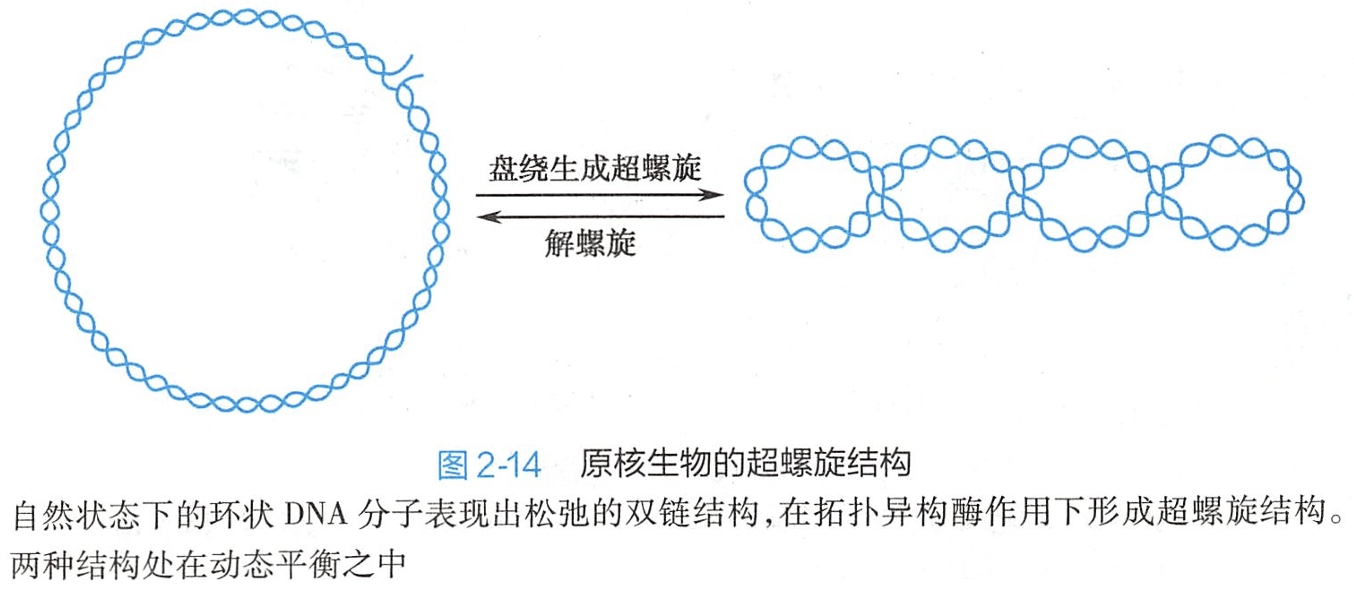
分析表明,在大肠杆菌的环状DNA中,平均每200个碱基就有一个负超螺旋形成。负超螺旋的DNA双链只能以封闭环状的形式或者在与蛋白质结合的条件下存在,以避免它们之间的相互纠缠。这种负超螺旋形式产生了DNA双链的局部解链效应,有助于诸如复制、转录等生物过程的进行。
线粒体和叶绿体是真核细胞中含有核外遗传物质的细胞器。,线粒体DNA(mitochondrial DNA, mtDNA)也是具有封闭环状的双螺旋结构。人mtDNA的长度是16569bp,编码了37个基因,包括13个蛋白质、2个rRNA和22个tRNA。
(二)真核生物DNA的高级结构
人类基因组大约有3×109个碱基对,这是一条长度约为1.7m的线性大分子。将这样的一条DNA双链组装在细胞核内,DNA双链需要进行一系列的盘绕、折叠和压缩。
在细胞周期的大部分时间里,细胞核内的DNA以松散的[200 学习/201 细胞生物学/第08章 细胞核/第2节 染色质和染色体/染色质和染色体|染色质]形式存在,只有在细胞分裂期间,细胞核内的DNA才形成高度致密的[200 学习/201 细胞生物学/第08章 细胞核/第2节 染色质和染色体/染色质和染色体|染色体]。在电子显微镜下观察到的染色质具有串珠样的结构(图2-15a)。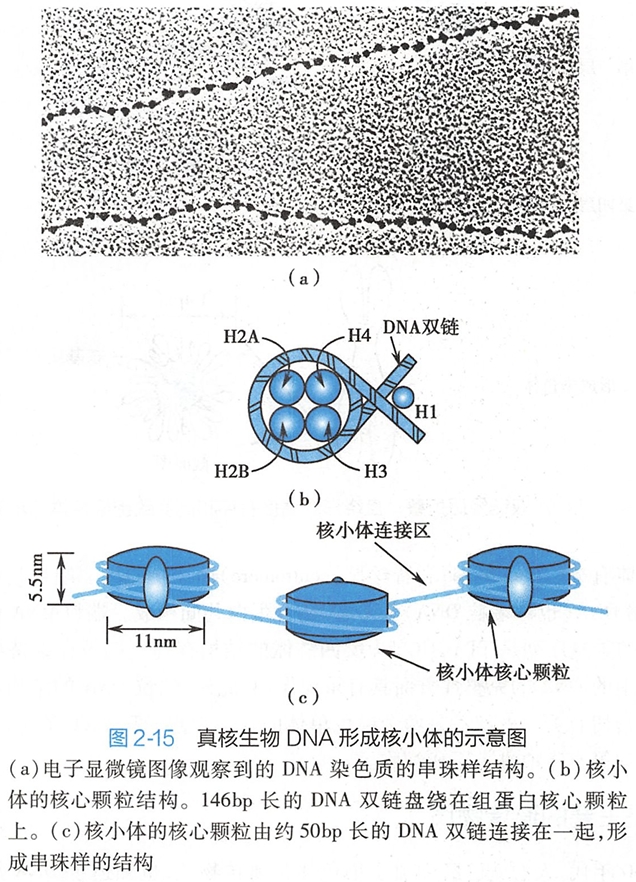
染色质基本组成单位是核小体(nucleosome),它是由一段双链DNA和4种碱性的组蛋白(histone,简写为H)共同构成的。
八个组蛋白分子(H2A×2,H2B×2,H3×2和H4×2)共同形成了一个八聚体的核心组蛋白,长度约146bp的DNA双链在核心组蛋白上盘绕1.75圈,形成核小体的核心颗粒(core particle)(图2-15b)。
核心颗粒是尺寸约11nmx6nm的盘状颗粒。连接相邻核小体之间的一段DNA称为连接段DNA(linker DNA),其长度在0~50bp之间不等,是非组蛋白结合的区域。组蛋白H1结合在盘绕在核心组蛋白上的DNA双链的进出口处,发挥稳定核小体结构的作用(图2-15c)。
至此,核小体核心颗粒和DNA双链形成了10nm的串珠状结构,也称为染色质纤维。这是DNA在核内形成致密结构的第一次折叠,使DNA的长度压缩了约7倍。
染色质纤维按照左手螺旋方式进一步盘绕卷曲,在组蛋白H1的参与下形成外径为30nm、内径为10nm的中空状螺线管(solenoidal)。
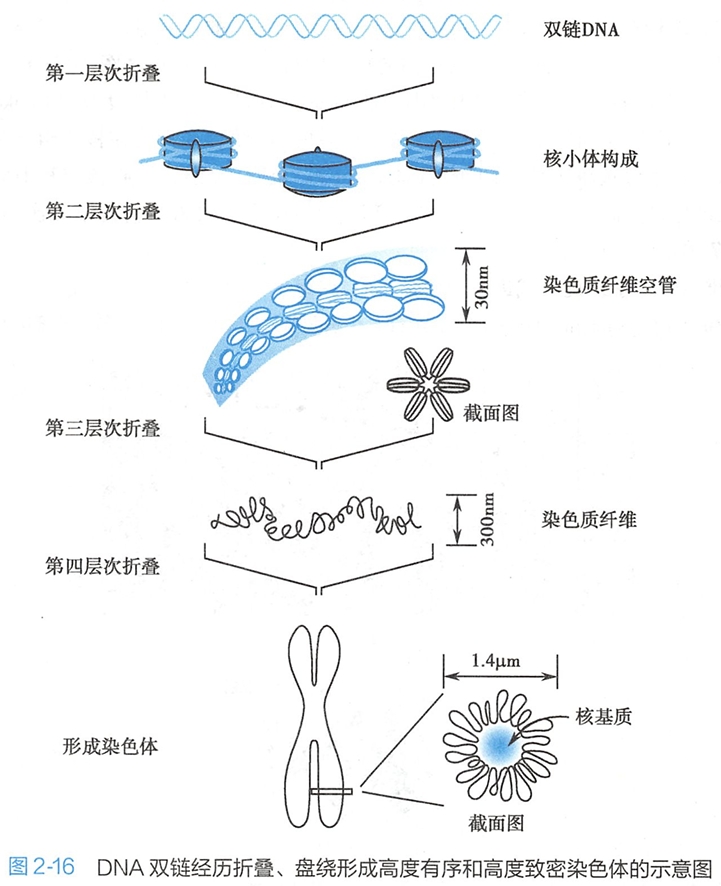
每个螺旋有6个核小体,组蛋白H1位于螺线管的内侧,继续发挥稳定螺线管的作用。染色质纤维中空状螺线管的形成与DNA特定区间的转录活性相关:正在进行转录的区间处在一种明显的无序状态之中,组蛋白质H1的数量也较少。染色质纤维中空状螺线管的形成是DNA在细胞内的第二次折叠,使DNA的压缩程度达到约40~60倍。
关于30nm中空状螺线管如何压缩成染色体,尚存争议。目前得到较为广泛认可的是多级螺线化模型(multiple coiling model)。染色质纤维螺线管的进一步卷曲和折叠形成了直径为400nm的超螺线管(supersolenoid),这一过程将DNA的长度又压缩了40倍。
之后,超螺线管的再度盘绕和压缩形成染色单体,在核内组装成染色体,使DNA长度又压缩了5~6倍(图2-16)。
这样,在染色体形成的过程中,DNA的长度总共被压缩了8000~10000倍,从而将近2m长的DNA有效地组装在直径只有几微米的细胞核中。
真核生物染色体有端粒(telomere)和着丝粒(centromere)两个功能区。
- 端粒是染色体端膨大的粒状结构,由染色体端DNA(也称端粒DNA)与DNA结合蛋白共同构成。端粒DNA由简单重复序列构成,人的端粒DNA的重复序列是TTAGGG,以G-四链体的结构存在。端粒在维持染色体结构的稳定性和维持复制过程中的DNA的完整性方面具有重要作用,此外,端粒DNA的结构和稳定性还与衰老及肿瘤的发生发展密切有关。
- 着丝粒是两个染色单体的连接位点,富含AT序列。细胞分裂时,着丝粒可分开使染色体均等有序地进入子代细胞。
三、DNA是主要的遗传物质
DNA是遗传信息的载体。生物体的遗传信息是以基因的形式存在的。
[200 学习/201 细胞生物学/第09章 基因表达/第1节 基因/基因|基因]是编码RNA或多肽链的DNA片段,即DNA中特定的核苷酸序列。
一个生物体的基因组(genome)是指包含在该生物的DNA(部分病毒除外)中的全部遗传信息,即一套染色体中的完整的核苷酸序列。各种生物体基因组的大小、所包含的基因数量和种类都有所不同。一般来讲,进化程度越高的生物体,其基因组越大越复杂。简单生物的基因组仅含有几千个碱基对,而高等动物的基因组可高达10碱基对,使可编码的信息量大大增加。
病毒颗粒的基因组可以由DNA组成,也可以由RNA组成,两者一般不共存。病毒基因组的DNA和RNA可以是单链的,也可以是双链的,可以是环形分子,也可以是线性分子。